Lecture 22: Exercise Physiology — Full Class Recap
35 slides
Slide 1
This final session is a course-wide recap that introduces no new material. It revisits the major conceptual themes of the course as a study guide for the final exam.
Students are encouraged to listen for unfamiliar points, note them, and return to the relevant earlier lecture for specifics — the goal is to synthesize knowledge around the recurring themes rather than memorize isolated facts.
Slide 2
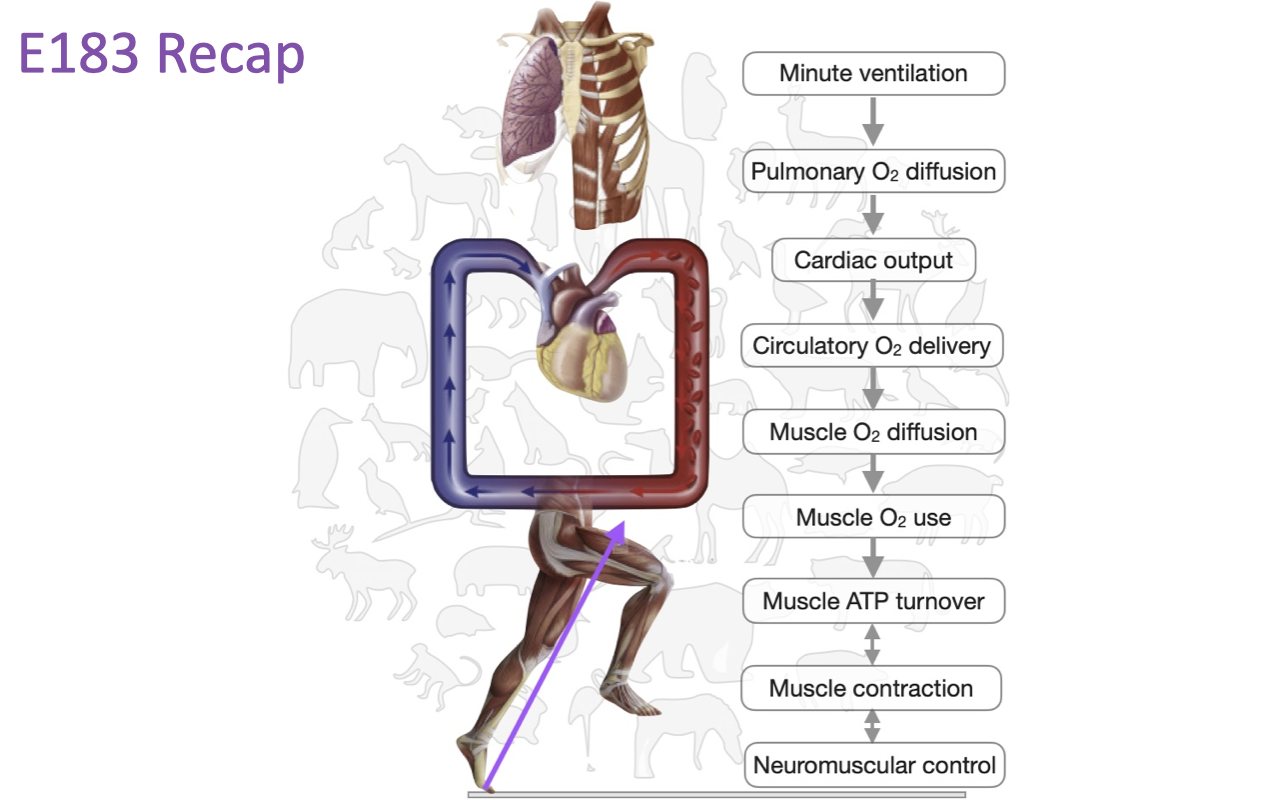
Course Overview: The Oxygen Cascade and Neuromuscular Control
The course examined the physiological systems and principles that determine and limit the ability to perform exercise, from oxygen supply through muscle contraction to neuromuscular control.
The focus is largely on humans but placed in a comparative and evolutionary context, which reveals where these systems show adaptability and plasticity versus specialization for particular functions.
Slide 3
Comparative Approaches: The Krogh Principle
The Krogh Principle holds that for many problems there is an ideal animal in which a physiological principle can be most conveniently studied (review Lecture 1).
Two comparative strategies recur through the course: case-study or model species (e.g., the cheetah for speed), and broader analysis of diversity and variation within evolutionary lineages.
Slide 4
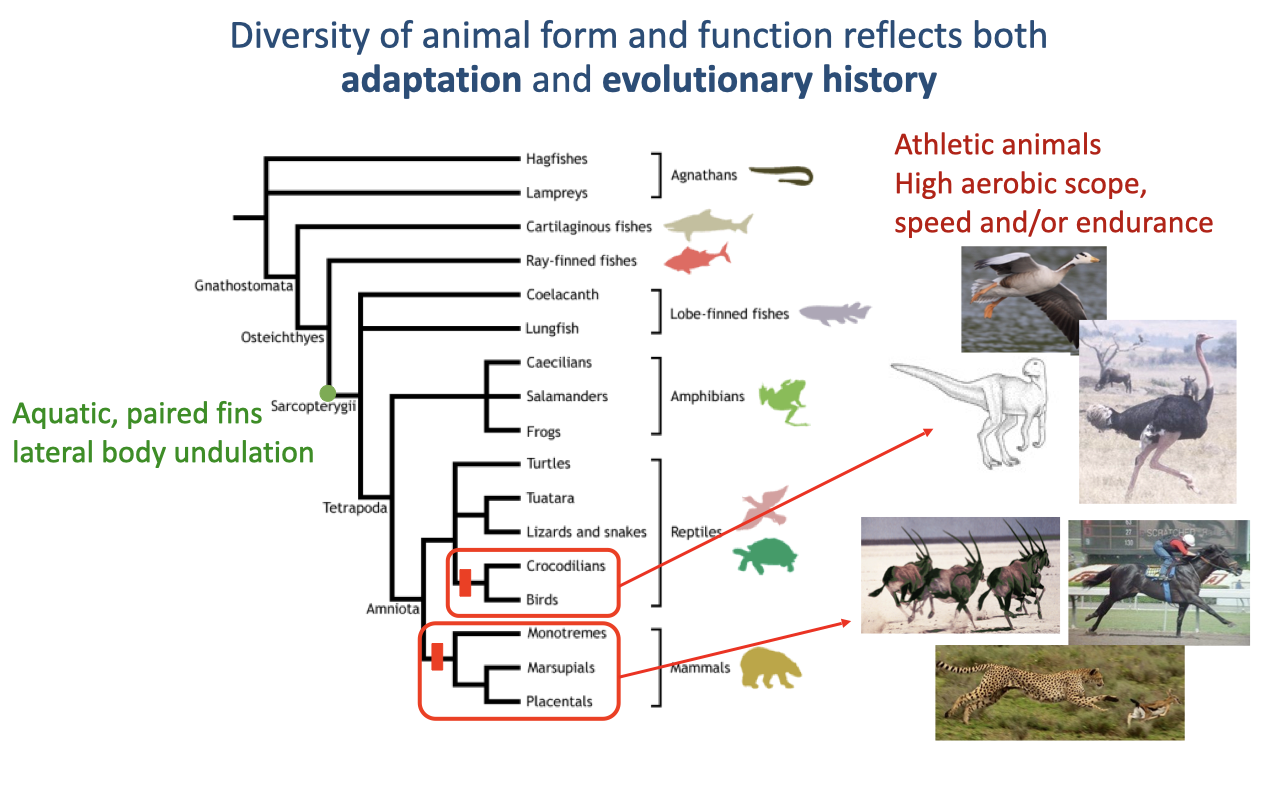
Diversity of Form Reflects Adaptation and Evolutionary History
Animal form and function reflect both adaptation and evolutionary history (review Lecture 1).
Vertebrates originated as aquatic animals; high-aerobic, athletic capacity evolved independently in two terrestrial lineages — mammals and the archosaurs (leading to dinosaurs and modern birds) — showing both convergent features and informative differences.
Slide 5
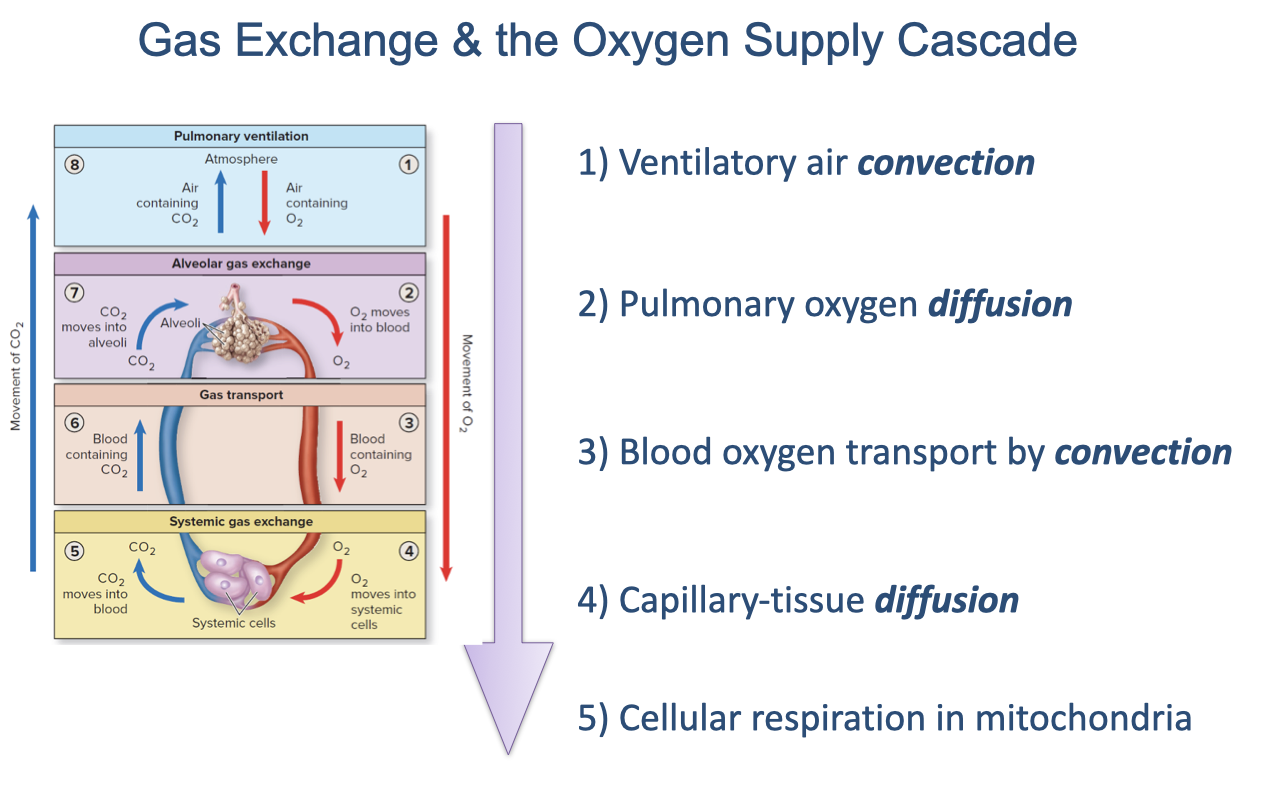
Gas Exchange and the Oxygen Supply Cascade
About four weeks covered the oxygen supply cascade — its sequential steps, governing equations, and limiting factors (review Lectures 1–10).
The five steps alternate convection and diffusion: ventilatory air convection, pulmonary diffusion, blood transport by convection, capillary-tissue diffusion, and mitochondrial respiration.
Partial pressure of oxygen falls at each step; mitochondrial O2 use is the sink that drives the cascade.
Slide 6
Two Key Equations: Alveolar Gas and the Fick Principle
The alveolar gas equation predicts alveolar oxygen from inspired oxygen, alveolar CO2, and the respiratory exchange ratio $R$ (typically 0.7–1.0); solving it requires the inspired fraction (~21%), atmospheric pressure (altitude-dependent), and water vapor pressure (~47 mmHg) (review Lecture 4).
The Fick principle calculates oxygen transport; the fractional-concentration form is preferred for exercise because ventilation and inspired/expired gas fractions can be measured non-invasively with a mask (review Lecture 4).
Slide 7
Changes and Limiting Factors in the Oxygen Cascade
The course examined how each cascade step changes, and what limits it, across five conditions (review Lectures 4–10).
These include exercise bouts in typical versus elite athletes, training adaptations (minute ventilation, stroke volume, heart rate), acute high-altitude responses in lowlanders, adaptations in high-altitude natives, and adaptations for acute hypoxia in diving.
Slide 8
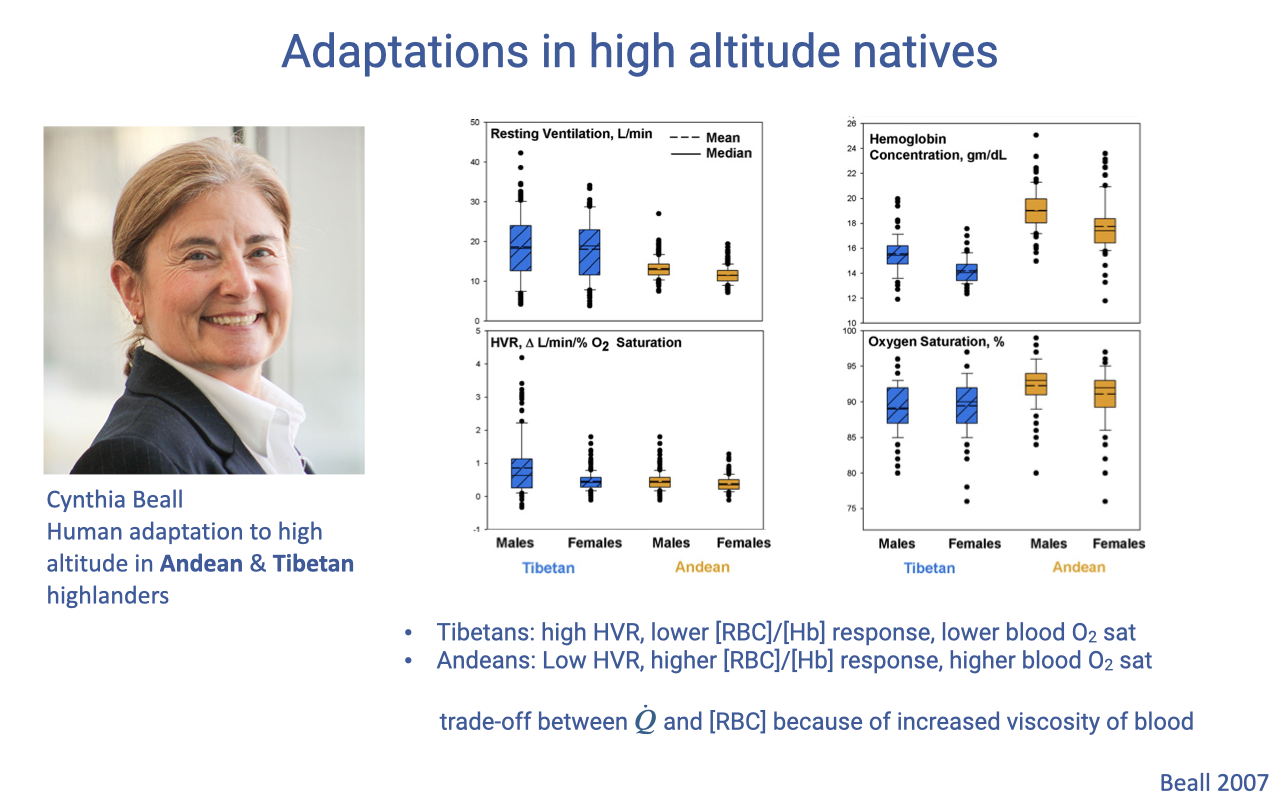
Adaptations in High-Altitude Natives: Tibetan vs. Andean
Different high-altitude human populations evolved different solutions to chronic hypoxia (review Lecture 9).
Tibetans show high ventilation and hypoxic ventilatory response but lower red cell count and saturation; Andeans show lower ventilation but higher red cell count and saturation.
A functional trade-off prevents combining both: high red cell count raises blood viscosity and cardiac work, and high ventilation demands high cardiac output for ventilation-perfusion matching — so each solution alone aids oxygen delivery and infant survival, but combined they are detrimental.
Slide 9
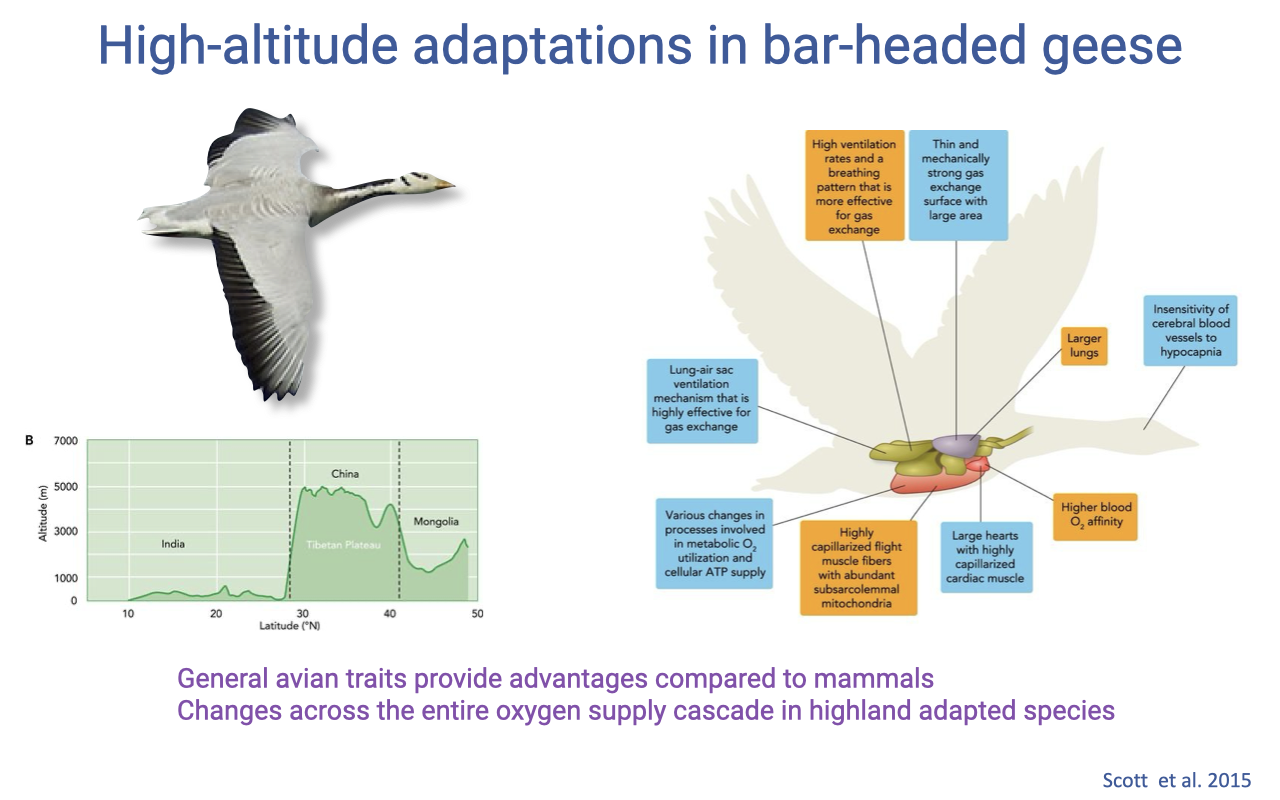
High-Altitude Adaptations in Bar-Headed Geese
Birds reach far higher altitudes than mammals, aided by general avian features — air sacs and a rigid lung that lowers the diffusion barrier without needing highly elastic, expandable lung tissue (review Lectures 6 and 9).
In highland-adapted species such as the bar-headed goose, advantageous changes occur across the entire oxygen supply cascade, not at one step alone.
Slide 10
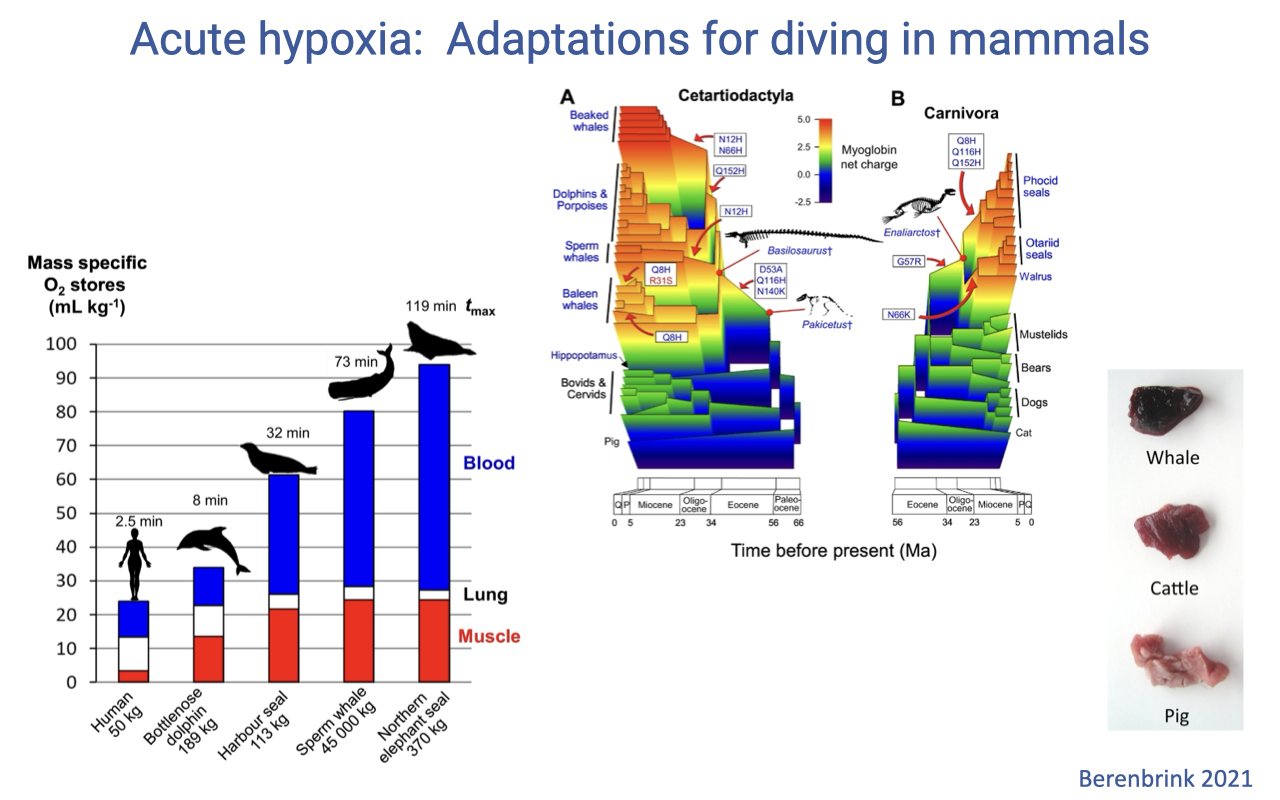
Acute Hypoxia: Adaptations for Diving in Mammals
Because ventilation is impossible underwater, diving performance depends on oxygen stored in the tissues rather than ongoing supply (review Lecture 10).
Divers have high mass-specific O2 stores, chiefly in blood (red blood cells, often released from large spleens during a dive) and in muscle (via myoglobin).
A surface-charge change in myoglobin, evolved independently several times, allows tighter myoglobin packing and the characteristically dark-red muscle of deep divers.
Slide 11
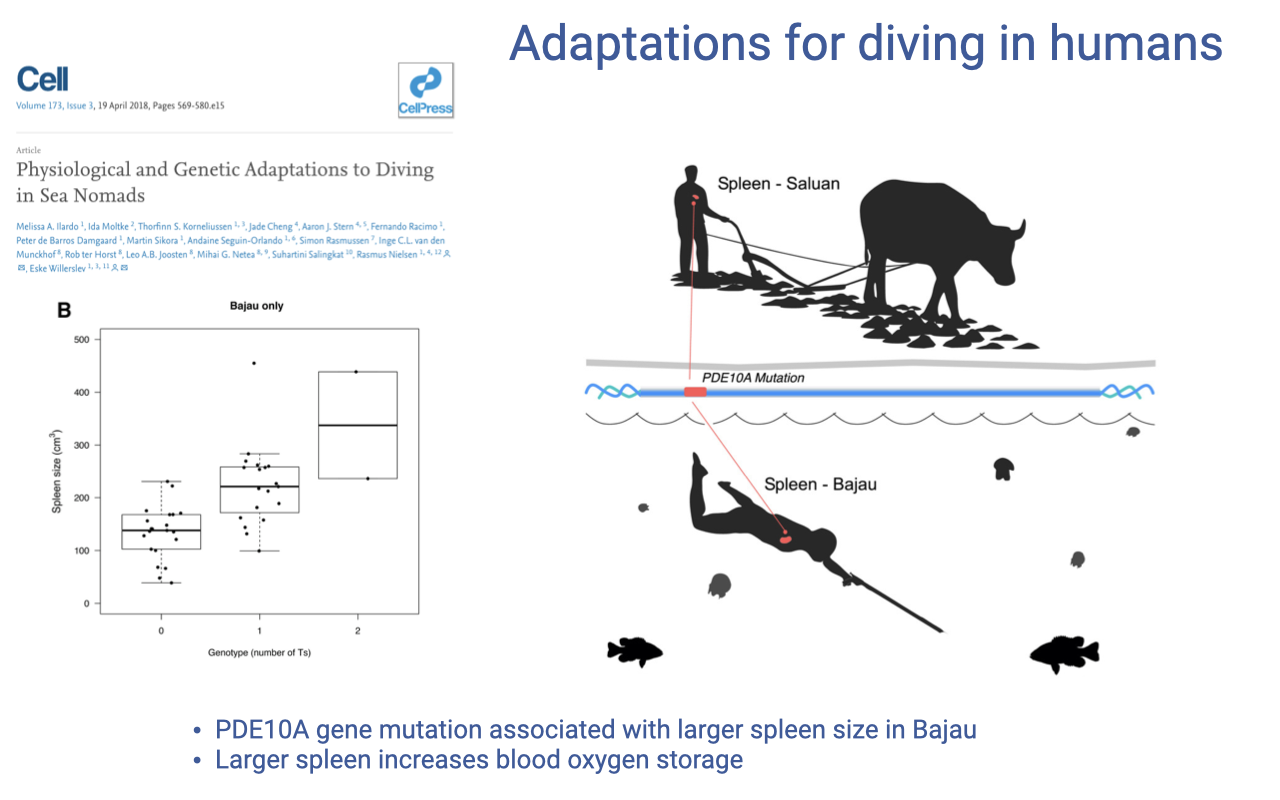
Adaptations for Diving in Humans: The Bajau
The Bajau “sea nomads” are persistent breath-hold divers who hold some of the longest human dive records (review Lecture 10).
A PDE10A gene variant in this population is associated with larger spleen size, providing extra red-cell oxygen storage that enhances diving performance.
Slide 12
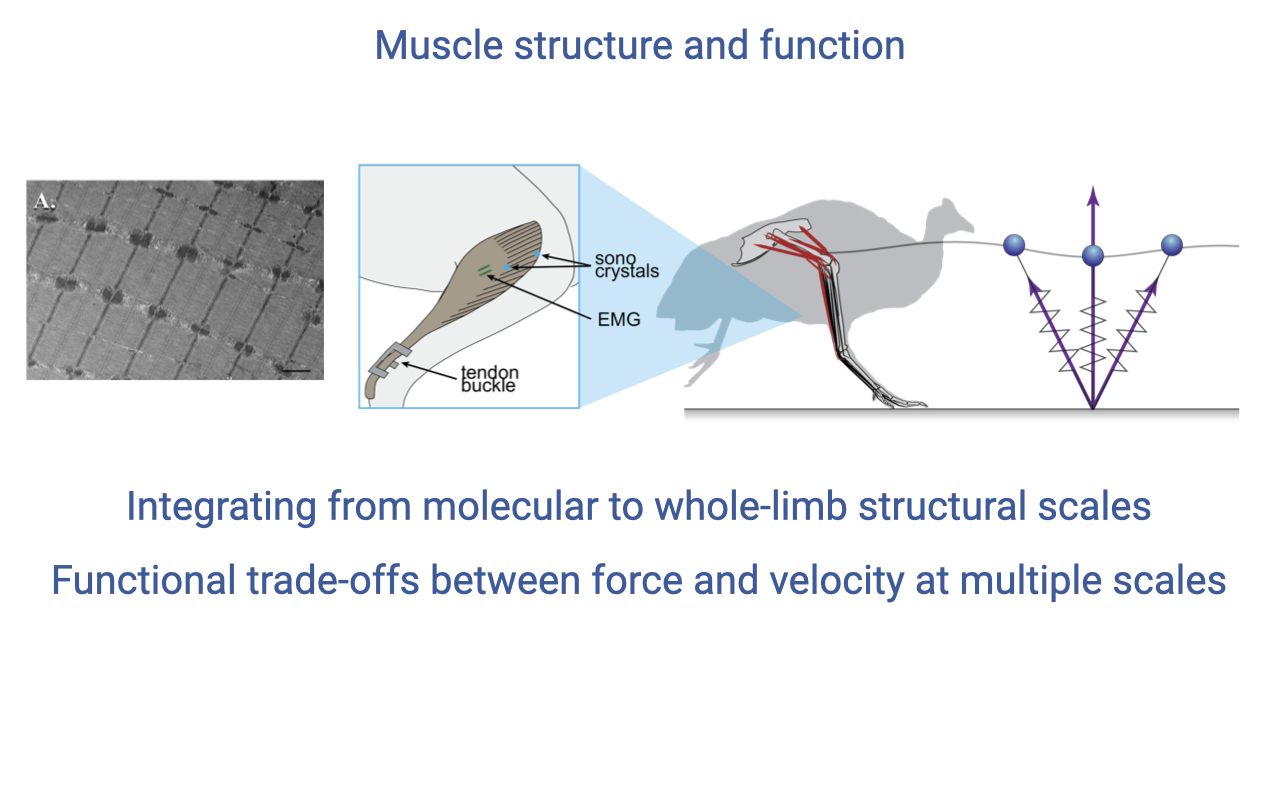
Transition: Muscle Structure and Function
The second half of the course shifted to muscle — the only motor in the system — progressing from molecular mechanisms up to whole-limb structure (review Lectures 11–18).
A unifying theme: force–velocity trade-offs appear at every scale and are additive, so each level contributes to whole-organism performance.
Slide 13
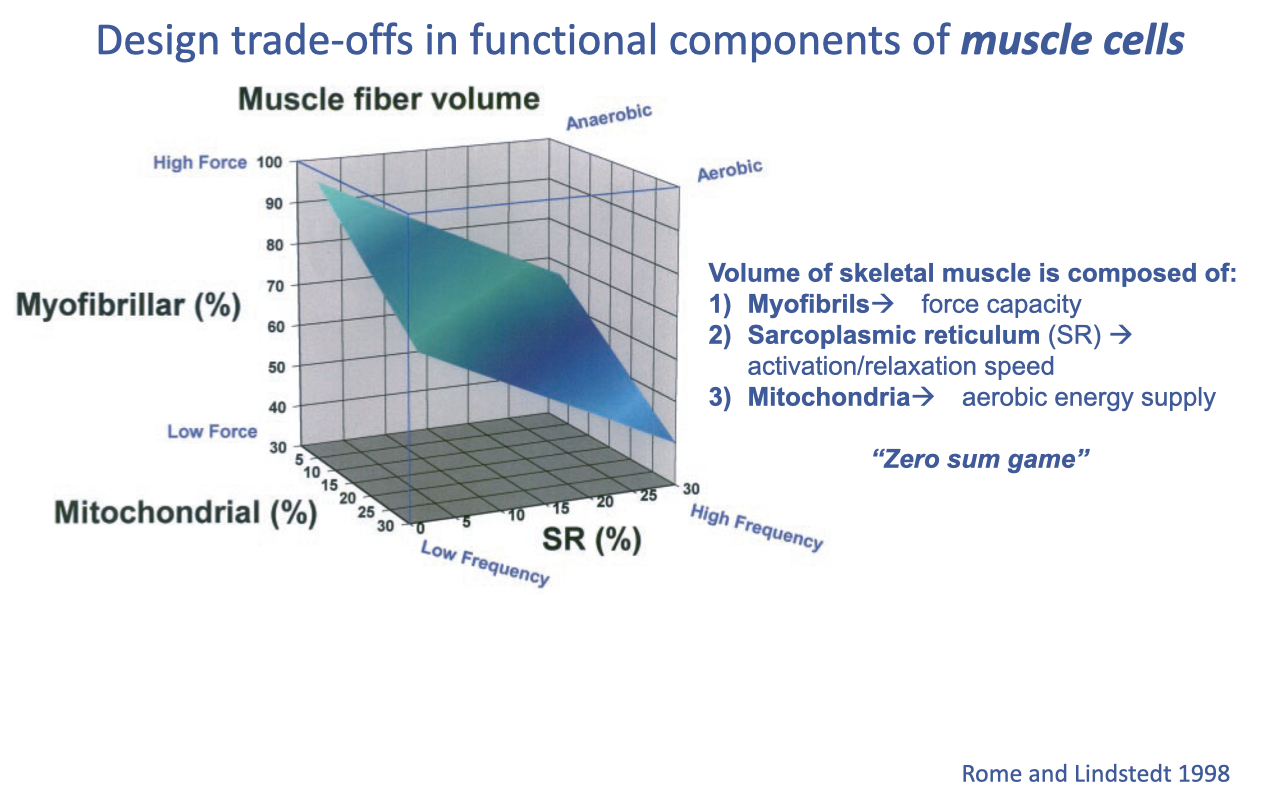
Design Trade-offs in Muscle Cells: The “Zero-Sum Game”
Inside a muscle cell, the volume fractions of myofibrils (force), sarcoplasmic reticulum (activation/relaxation speed), and mitochondria (aerobic capacity) compete — a “zero-sum game” for cell volume (review Lecture 11).
This is zero-sum only for volume allocation, not total size — hypertrophy still raises force by adding bulk. The trade-off explains why aerobic muscles, packed with mitochondria, generate lower force, underlying the strength-versus-endurance trade-off.
Slide 14
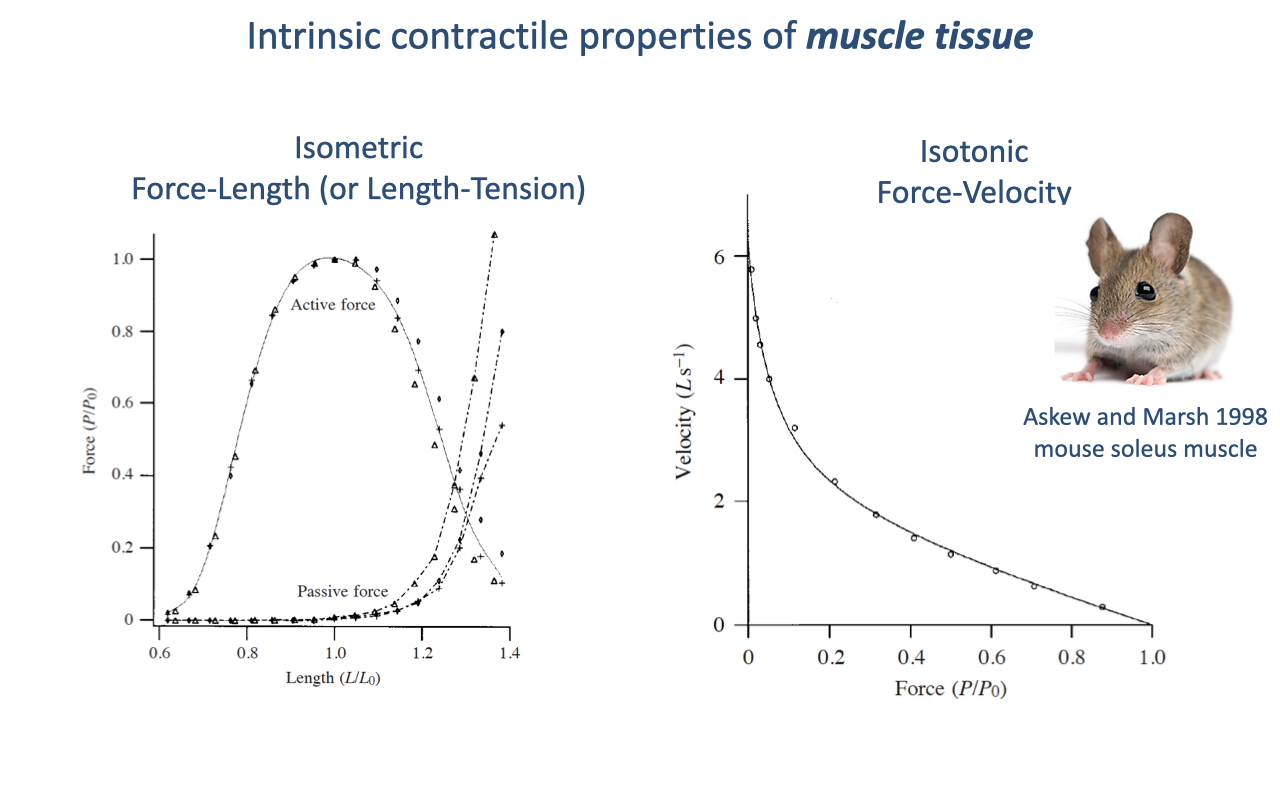
Intrinsic Contractile Properties of Muscle Tissue
The force–length relationship sets a performance envelope: peak force occurs near optimal length and falls at shorter or longer lengths, so muscles generate force over only a limited excursion (review Lecture 12).
The force–velocity relationship shows an inherent force-versus-speed trade-off; because power is force times velocity, peak power occurs at intermediate contraction speeds.
Slide 15
Comparative Maximum Velocity and Body Size
Maximum (unloaded) shortening velocity differs by fiber type — fast versus slow — and also scales with body size (review Lecture 12).
Smaller animals have faster muscles; larger animals have slower muscles, partly because larger animals are constrained to trade speed for the force needed to support body weight.
Slide 16
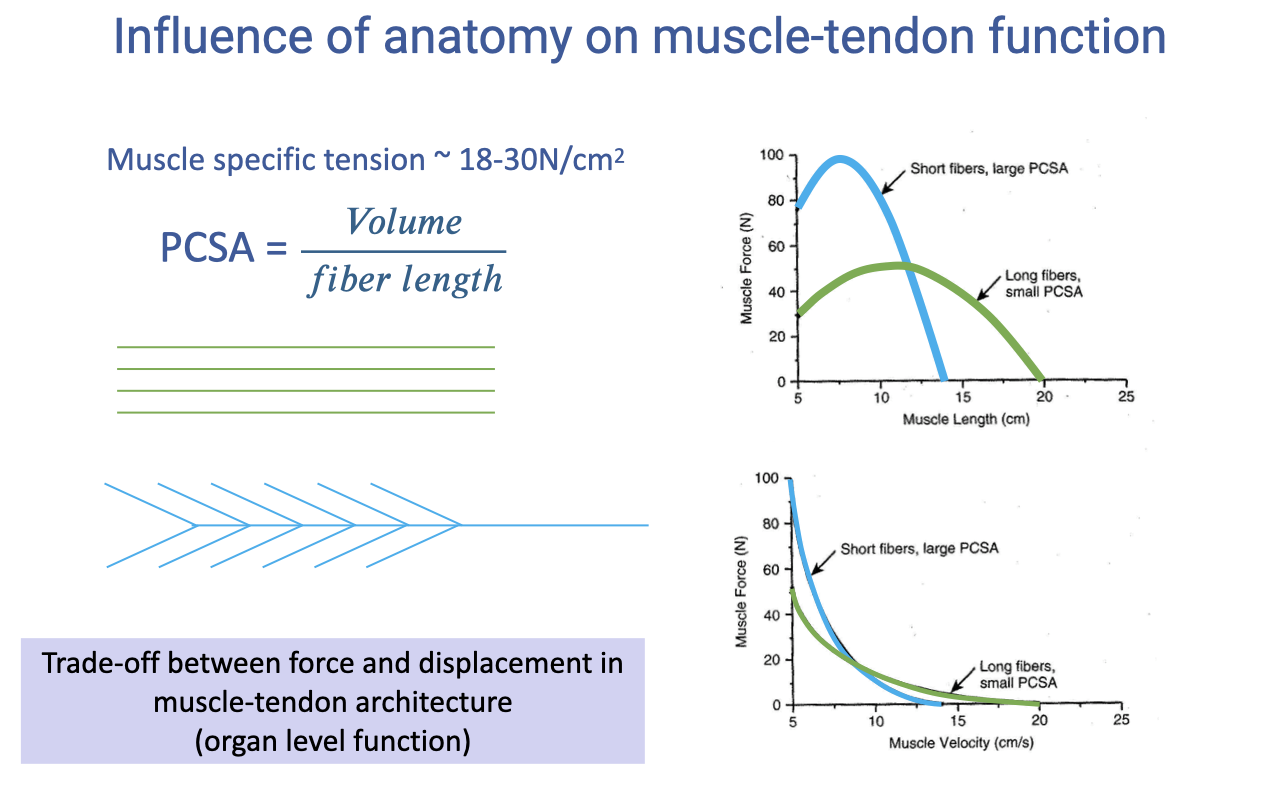
Influence of Anatomy on Muscle-Tendon Function
Specific tension (~18–30 N/cm²) is fairly conserved, so physiological cross-sectional area (PCSA) — volume divided by fiber length — sets a muscle’s force capacity (review Lecture 13).
For equal volume, pennate (short-fiber) muscles produce higher force but less displacement, while parallel (long-fiber) muscles produce more displacement and velocity — an organ-level force-versus-displacement trade-off.
Slide 17
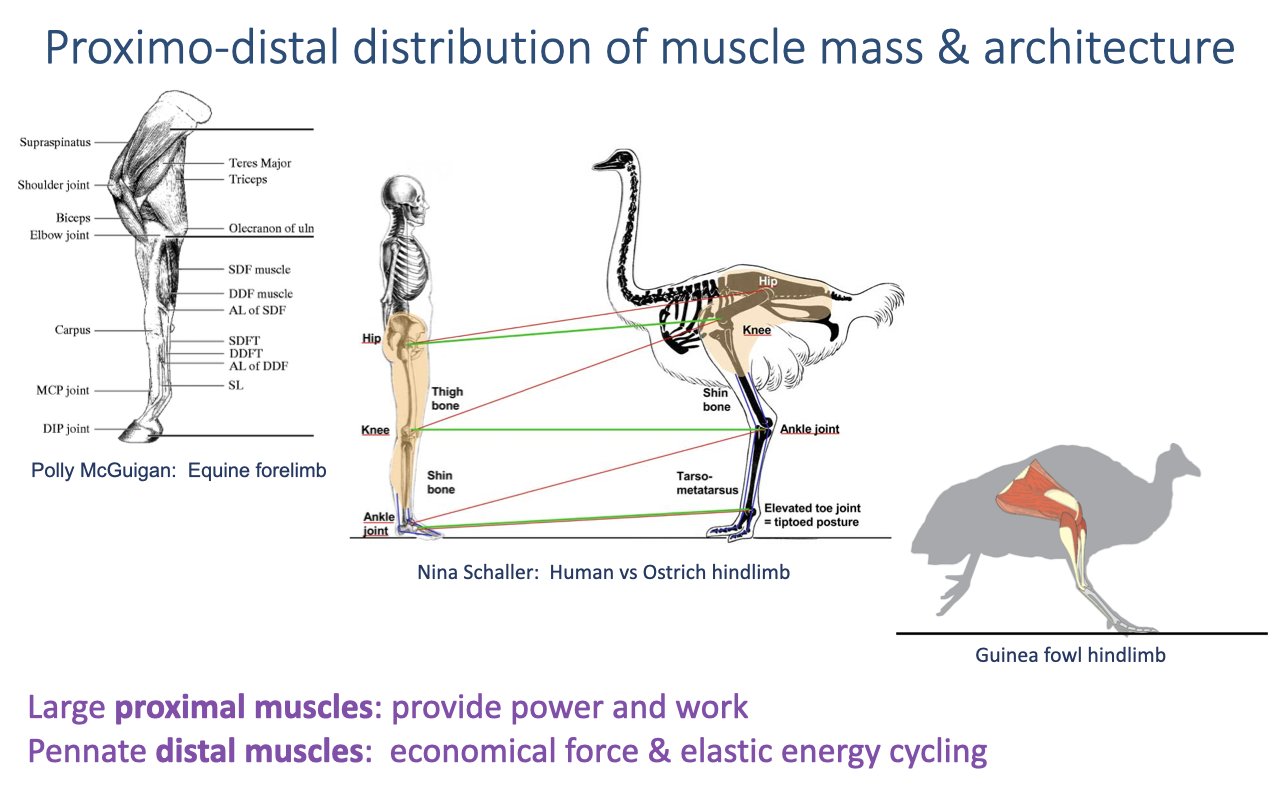
Proximo-Distal Distribution of Muscle Mass
Limbs distribute muscle architecture by joint position rather than using one generalist design (review Lectures 13–14).
Large proximal muscles (hip, knee) have parallel fibers and large volume for high work and power; pennate distal muscles (e.g., calf) give economical force and elastic energy cycling.
Keeping distal limbs light reduces limb inertia and the energy cost of swinging the leg.
Slide 18
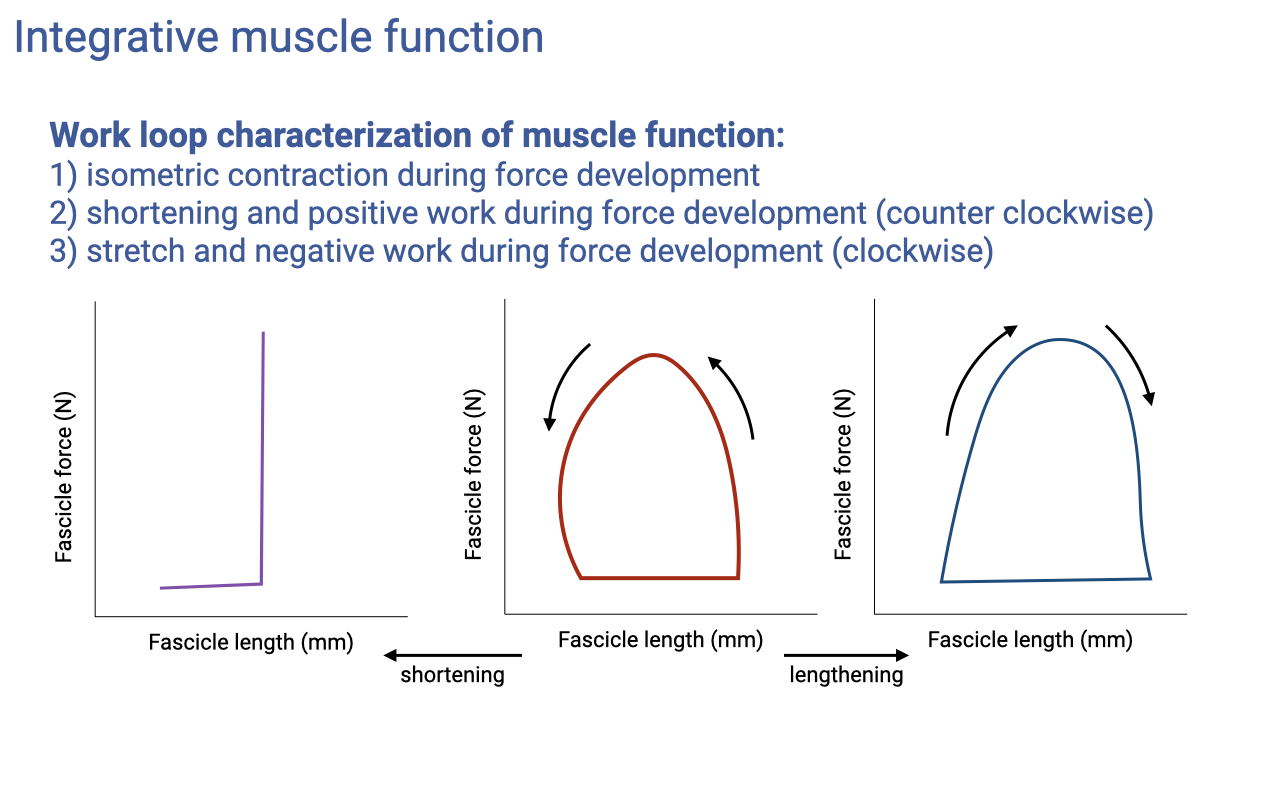
Integrative Muscle Function: Work Loops
Work loops classify muscle action by the shape traced in force–length space (review Lecture 14).
Isometric (strut-like, near-vertical) muscles let springy tendons cycle elastic energy; counter-clockwise loops do positive work (motor-like, e.g., bird pectoralis, human hip); clockwise loops absorb energy (brake-like, e.g., landing from a jump).
Slide 19
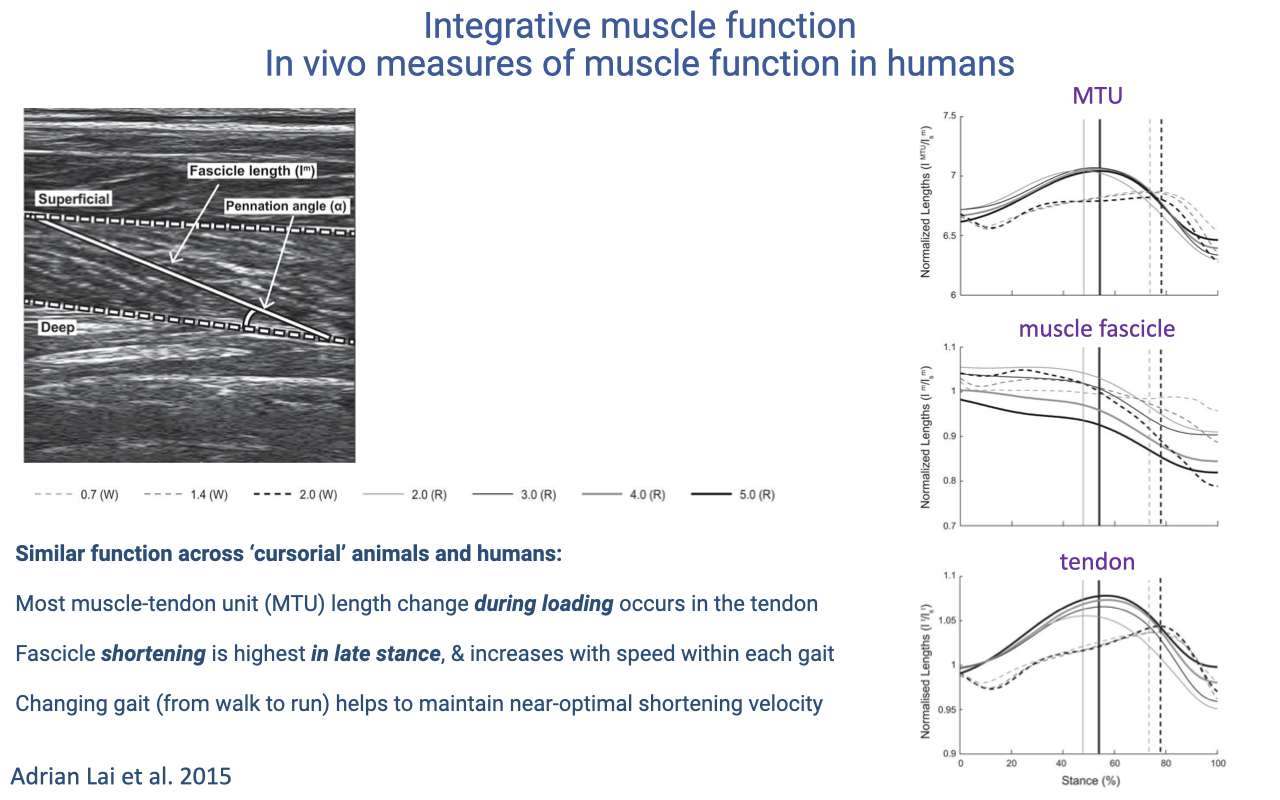
In Vivo Muscle Function in Humans
Ultrasound studies of the human calf reveal that during the first half of stance the tendon stretches while fascicles stay near-isometric; positive fascicle shortening occurs mostly in late stance to re-accelerate the body (review Lecture 14).
Fascicle shortening rises with speed within a gait; switching from walking to running keeps fascicle velocity in a more efficient range by shifting load and stretch into the elastic tendons.
Slide 20
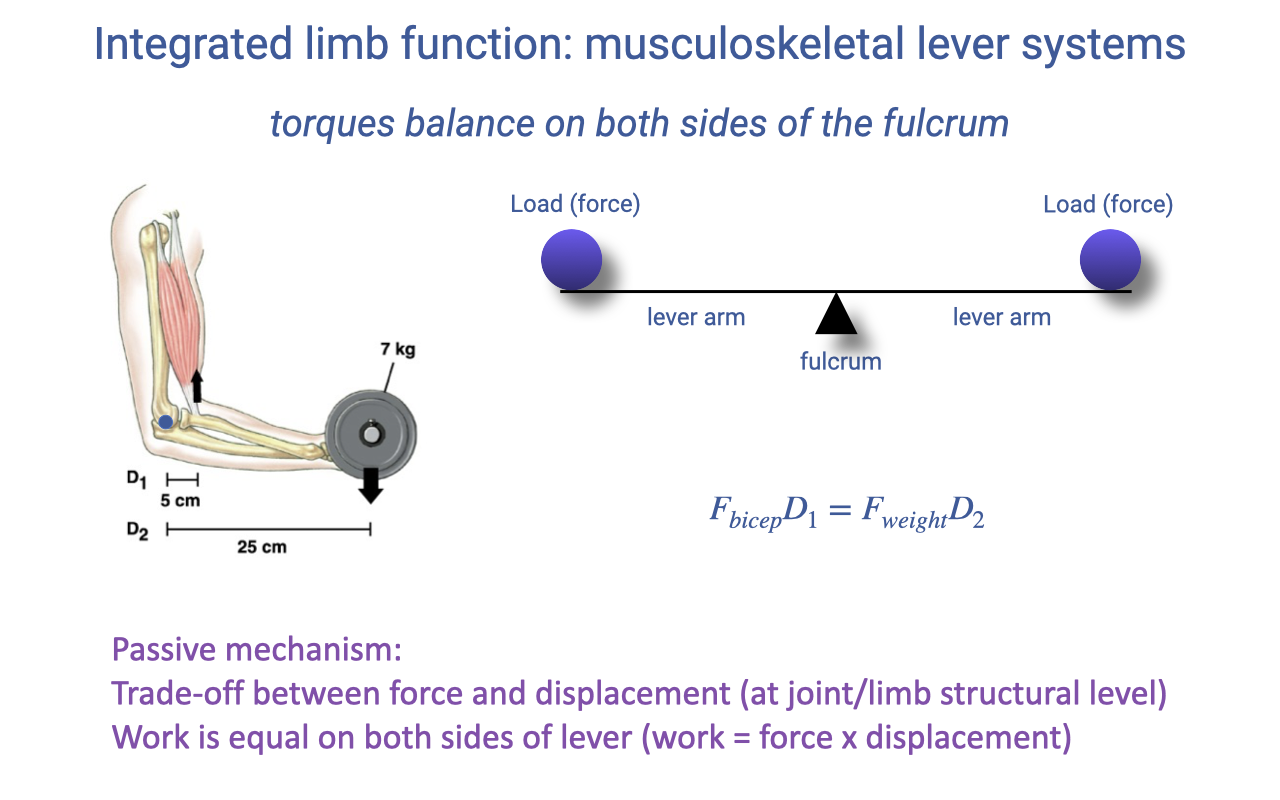
Integrated Limb Function: Musculoskeletal Lever Systems
A lever system balances torques across the fulcrum: muscle force times its moment arm equals load force times its moment arm (review Lecture 14).
As a passive mechanism it trades force for displacement but conserves total work — a high-force, low-excursion arrangement versus a low-force, high-excursion one.
Slide 21
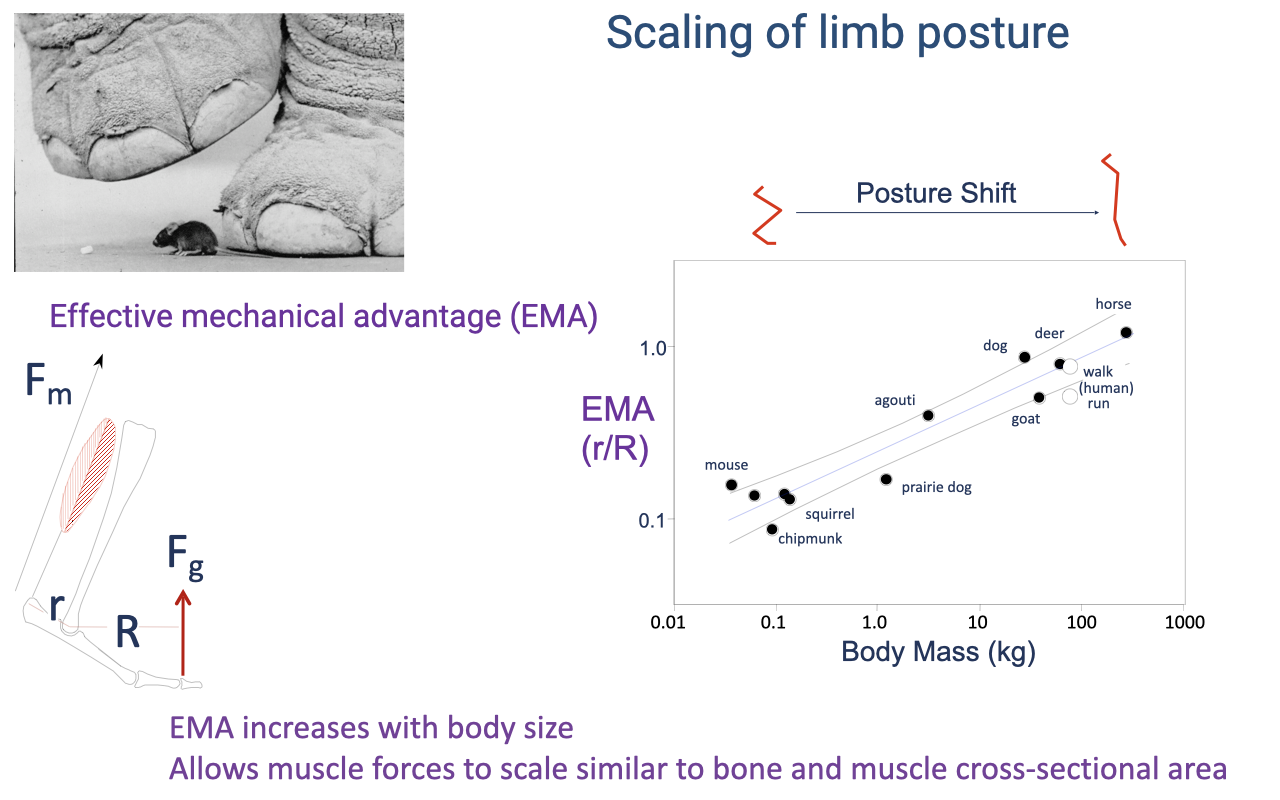
Scaling of Limb Posture
Across body size there is a posture shift from crouched (flexed) limbs in small animals to straighter limbs in large animals (review Lecture 14).
This raises effective mechanical advantage (EMA) with size, letting muscle force scale in step with bone and muscle cross-sectional area so large animals can support their weight against gravity.
Slide 22
Principles of Training
Four training principles govern adaptation (review Lectures 15 and 21): overload (stress beyond the usual), progression (gradually increasing load), specificity (adaptations match the system and movement trained), and reversibility (detraining when training stops).
Different forms of activity produce training responses on different timescales, and detraining likewise proceeds at different rates for different aspects of fitness — for example, VO2 and strength are lost at different rates.
Slide 23
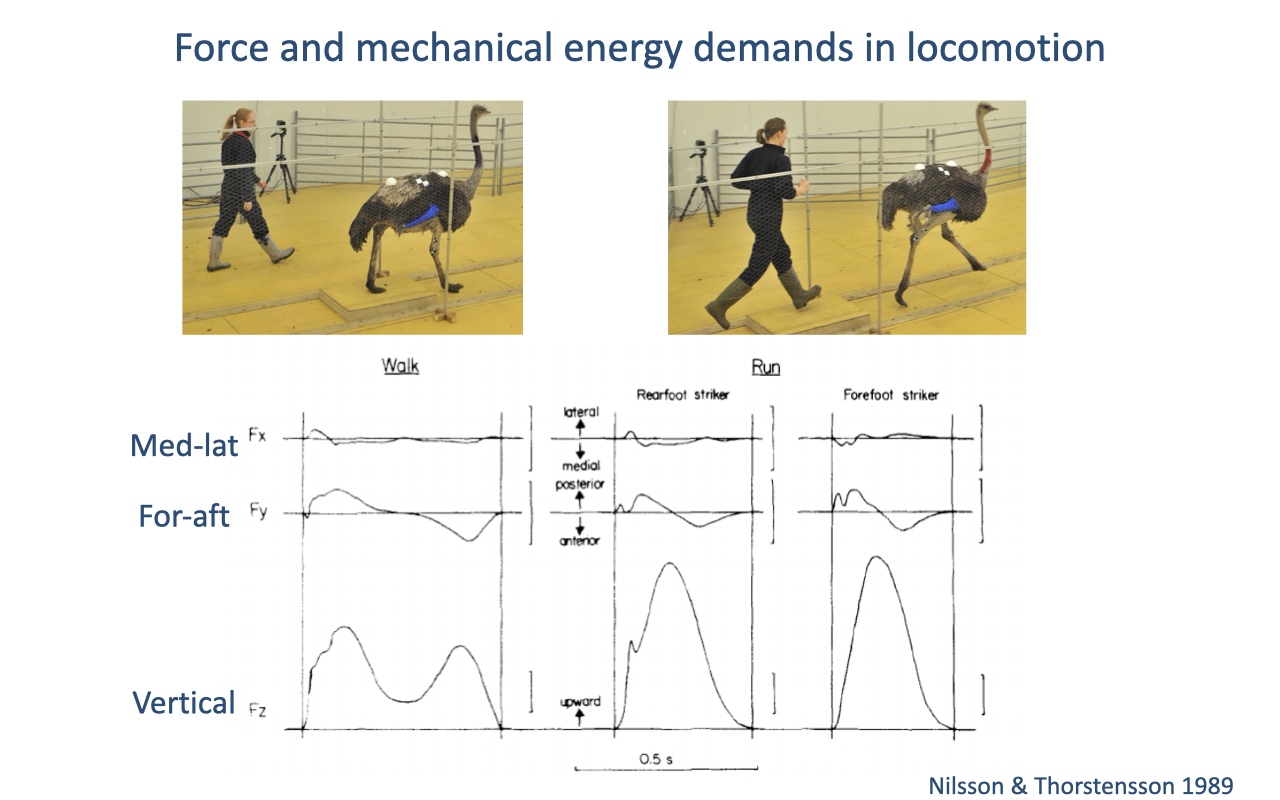
Force and Mechanical Energy Demands in Locomotion
Ground reaction forces have vertical, fore-aft (braking then propulsive), and mediolateral components that vary with gait (review Lectures 16–17).
Ground reaction force demands determine the muscle forces required, and therefore the effort and energetic cost of locomotion.
Slide 24
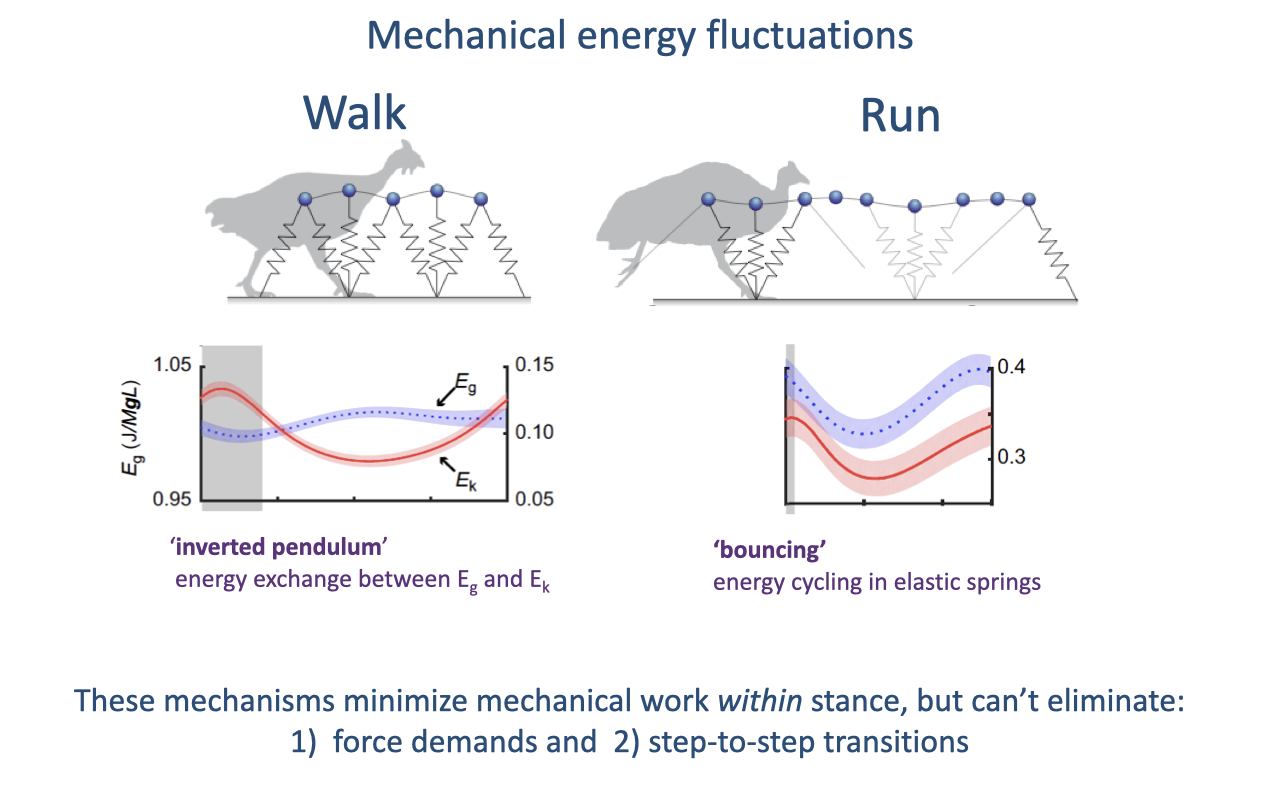
Mechanical Energy Fluctuations: Walking vs. Running
Walking uses inverted-pendulum exchange between gravitational potential and kinetic energy (out of phase); running uses elastic bouncing where the two energies fluctuate in phase (review Lecture 17).
Both passive mechanisms reduce the mechanical work of locomotion but cannot eliminate the force demands of supporting body weight or the collisional energy losses of step-to-step transitions.
Slide 25
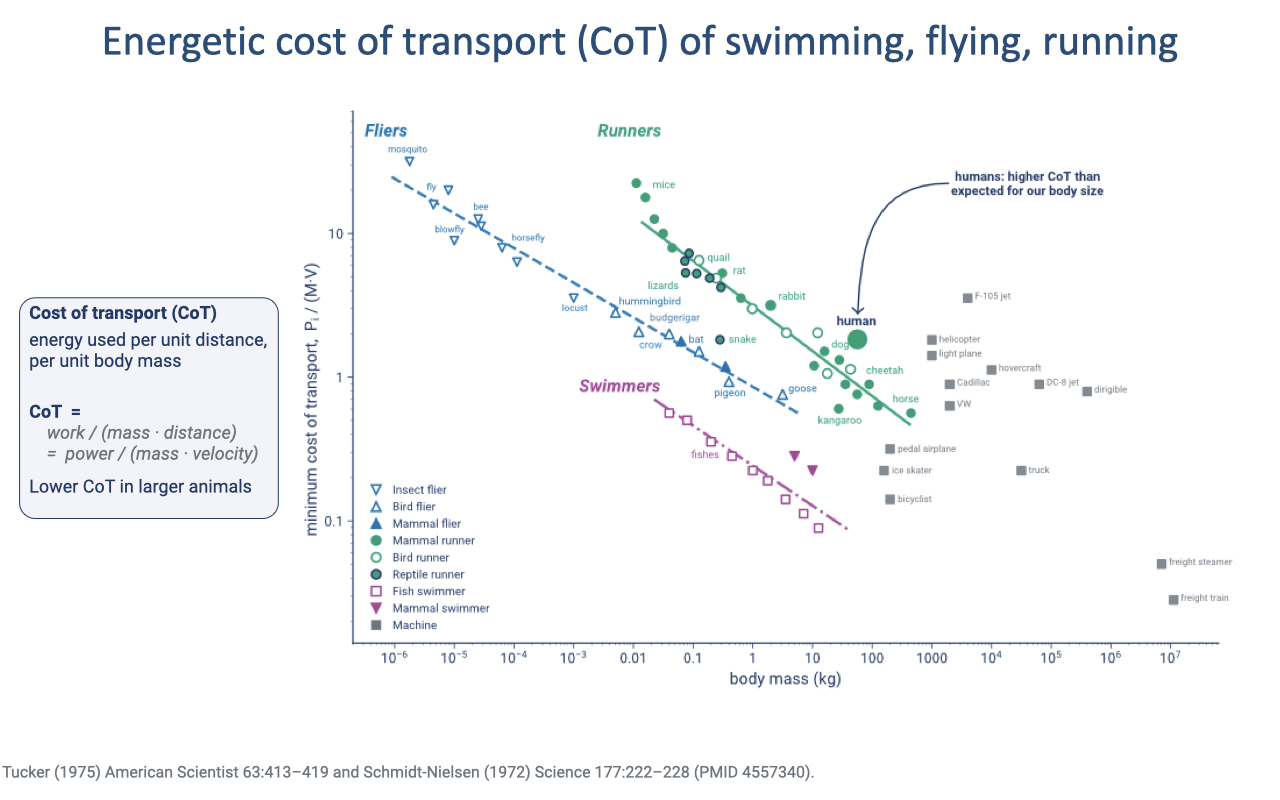
Energetic Cost of Transport
Cost of transport (CoT) is energy used per unit distance per unit body mass; for a given mode it declines with body size — larger animals move more economically per kilogram (review Lecture 18).
Running is most costly, flying intermediate, and swimming cheapest, because the force demands differ by mode: runners support body weight against gravity and suffer collisional losses at each foot contact, fliers support weight without collisional losses, and swimmers need not support weight at all.
Humans sit somewhat above the expected running line for their size.
Slide 26
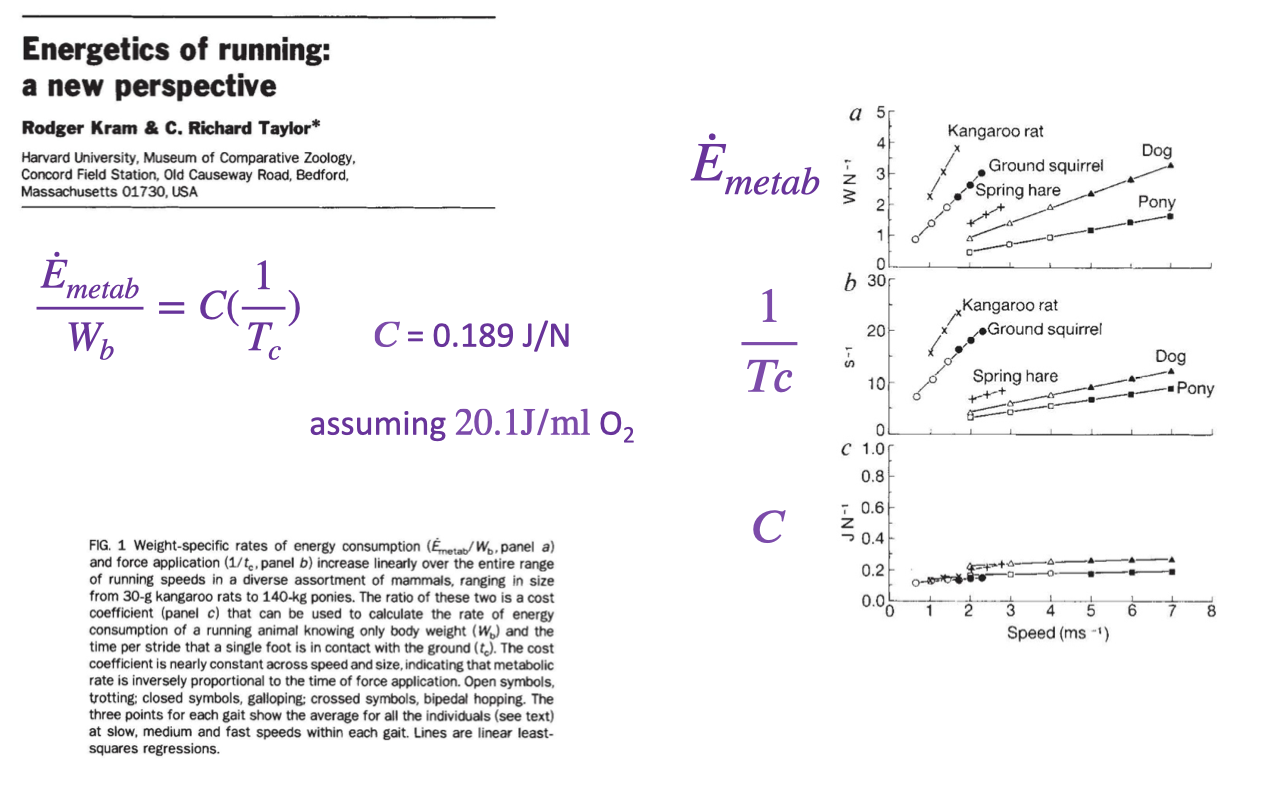
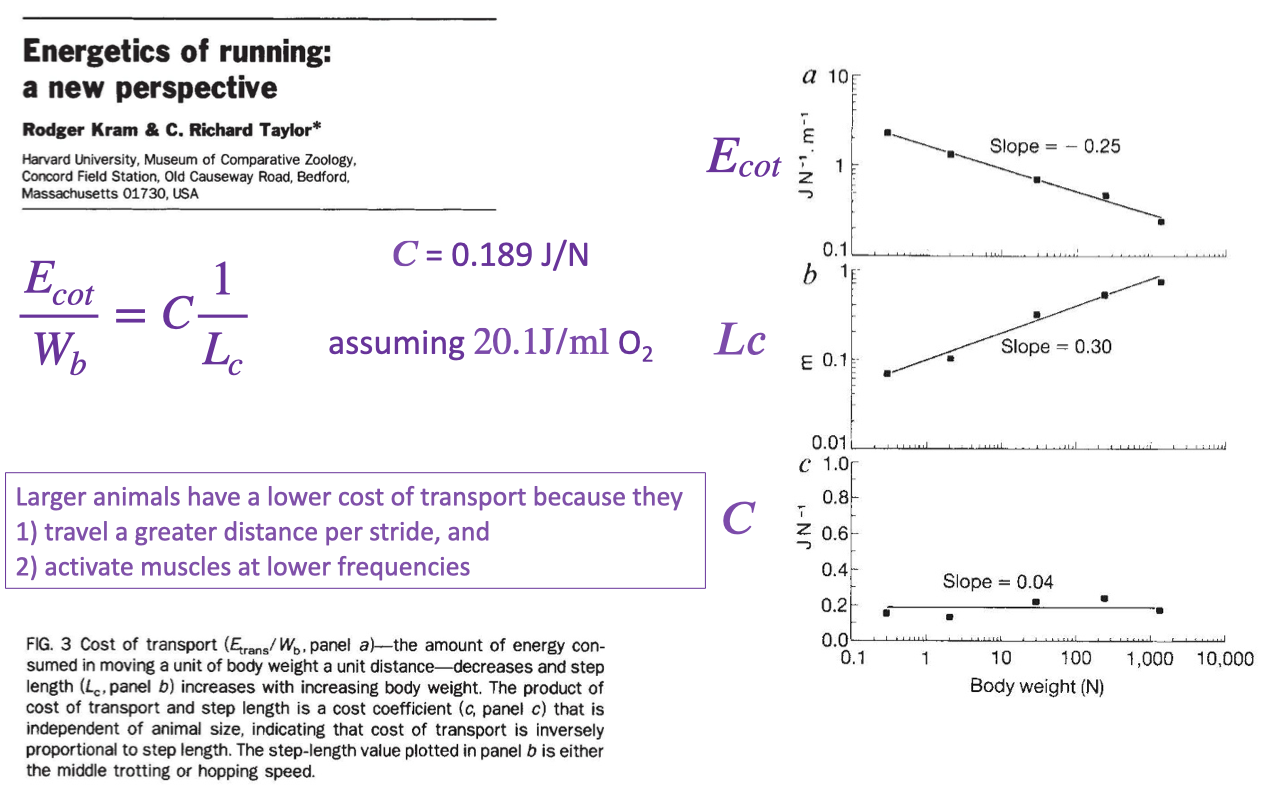
Energetics of Running: A Cost-Coefficient View
Kram and Taylor showed that running metabolic rate scales with the rate of force generation — the inverse of foot-ground contact time, $1/T_c$ (review Lecture 18).
A nearly constant cost coefficient (~0.189 J/N) links the energy cost to how quickly the limbs must be cycled to apply support force.
Slide 27
Why Larger Animals Have Lower Cost of Transport
Larger animals have lower cost of transport because they cover more distance per stride and turn their muscles on and off at lower frequencies, giving longer ground-contact times and lower force-generation costs (review Lecture 18).
The cost coefficient stays roughly constant across body size, and the same scaling has been replicated within humans of differing stature.
Slide 28
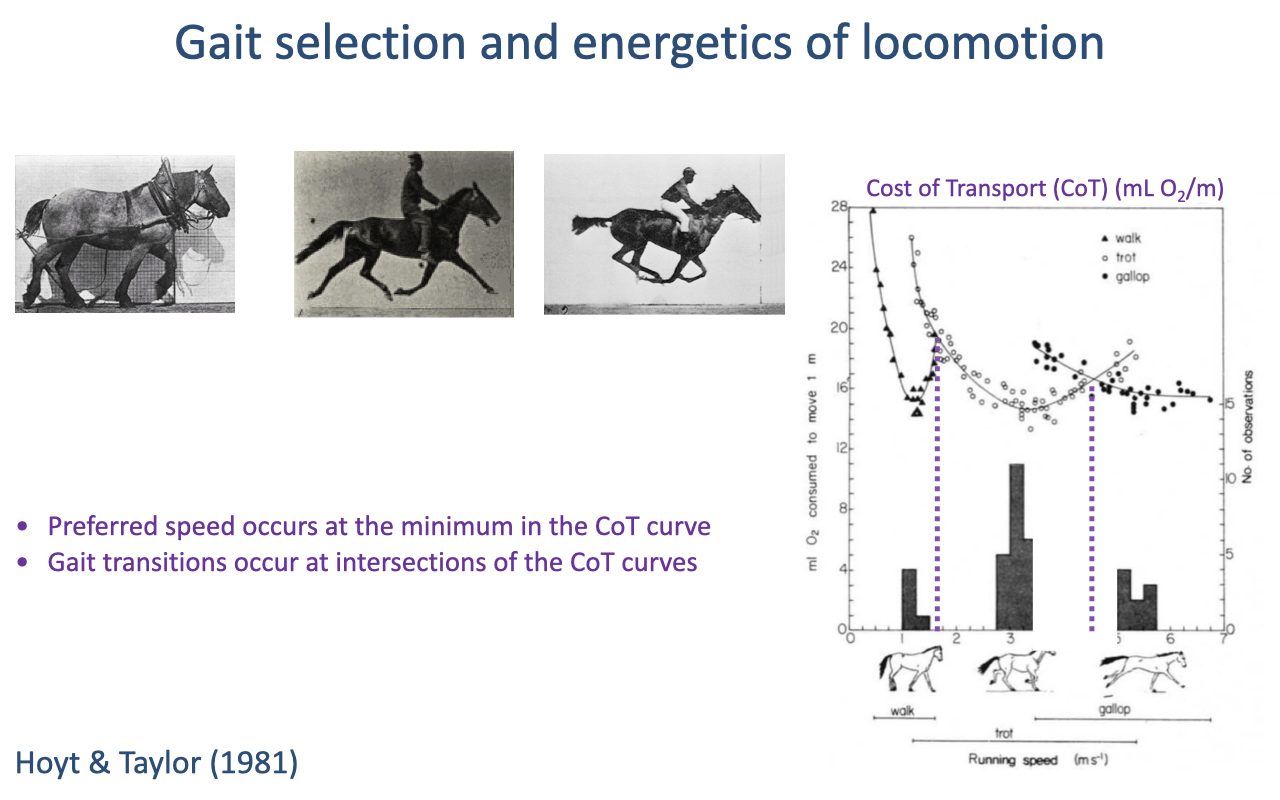
Gait Selection and the Energetics of Locomotion
Within a gait, cost of transport follows a U-shaped curve when animals are forced off their preferred speed — moving faster or slower than preferred both raise the cost (review Lecture 18).
Preferred speeds occur at each curve’s minimum, and gait transitions occur where adjacent curves intersect, which requires projecting the curves to locate the crossing points.
Slide 29
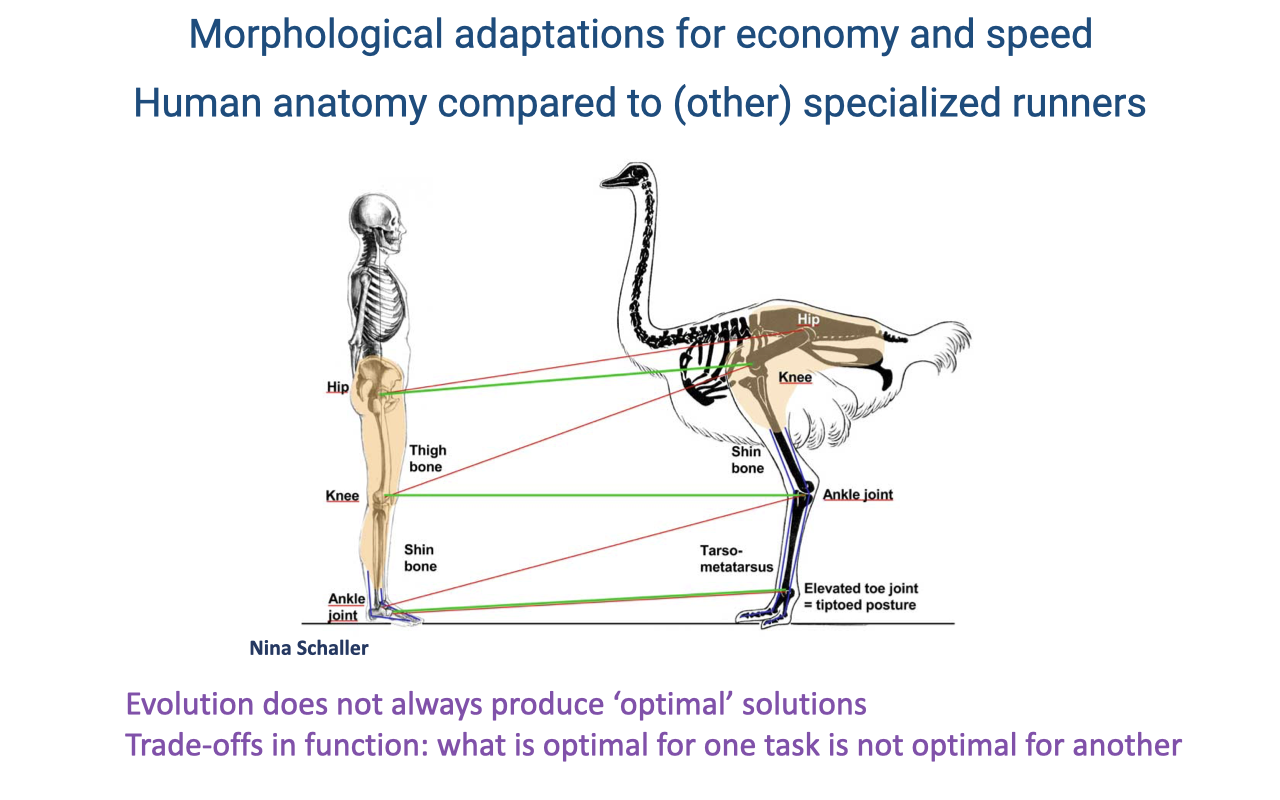
Morphological Adaptations for Economy and Speed
Athletic demands differ: migrating long distances favors economy, while escaping predators favors speed — some features serve both, but not all (review Lecture 18).
Whether humans count as specialized runners is debated. Humans have long legs (suggesting athletic specialization) but heavy, massive legs compared with specialized runners such as the ostrich.
Evolution does not always produce optimal solutions — it tinkers with inherited form, so human function must be read against our primate ancestry, and trade-offs mean what is optimal for running need not be optimal for walking or non-locomotor tasks.
Slide 30
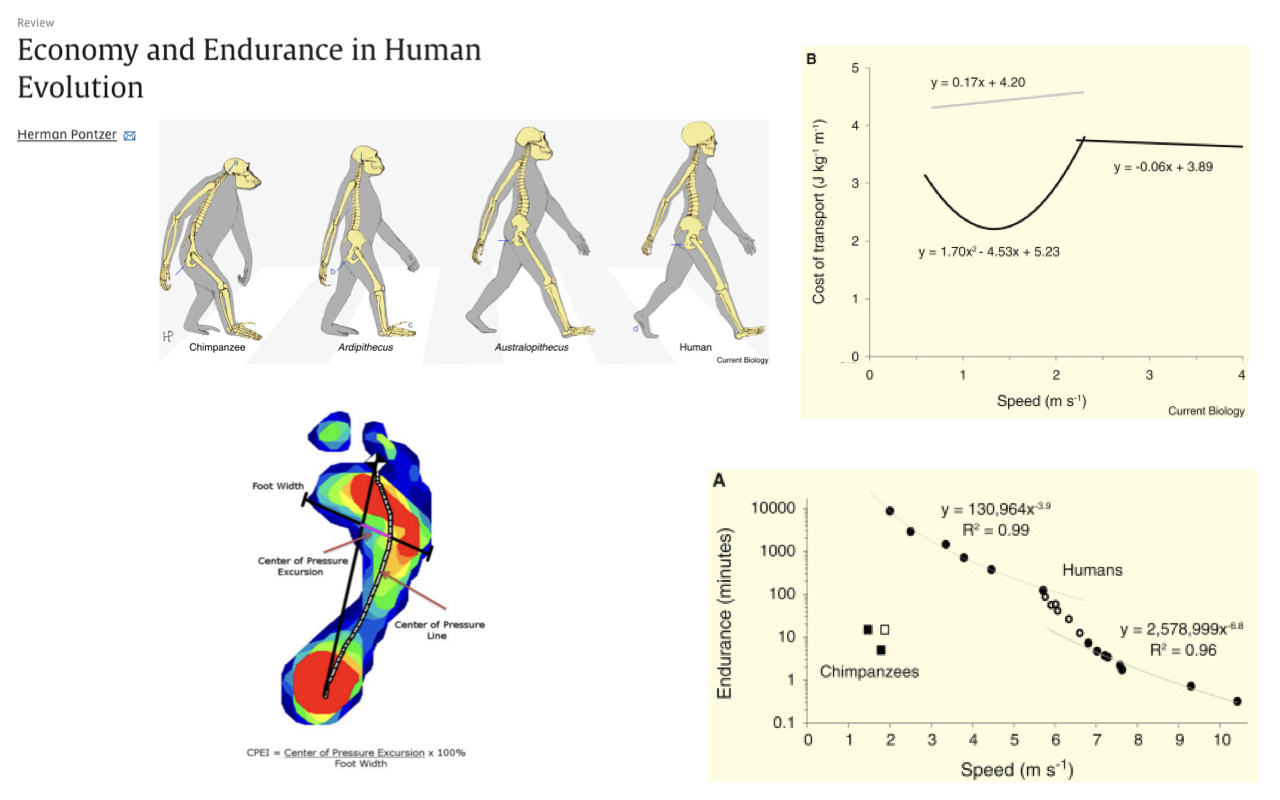
Economy and Endurance in Human Evolution
Placed against great-ape ancestry, humans have exceptionally economical walking — a much lower cost of transport than chimpanzees, with a clear U-shaped optimum (review Lecture 18).
Humans also show exceptional endurance in both walking and running, along with physiological specializations such as heat dissipation that support sustained activity — consistent with selection for economical, enduring movement.
Slide 31
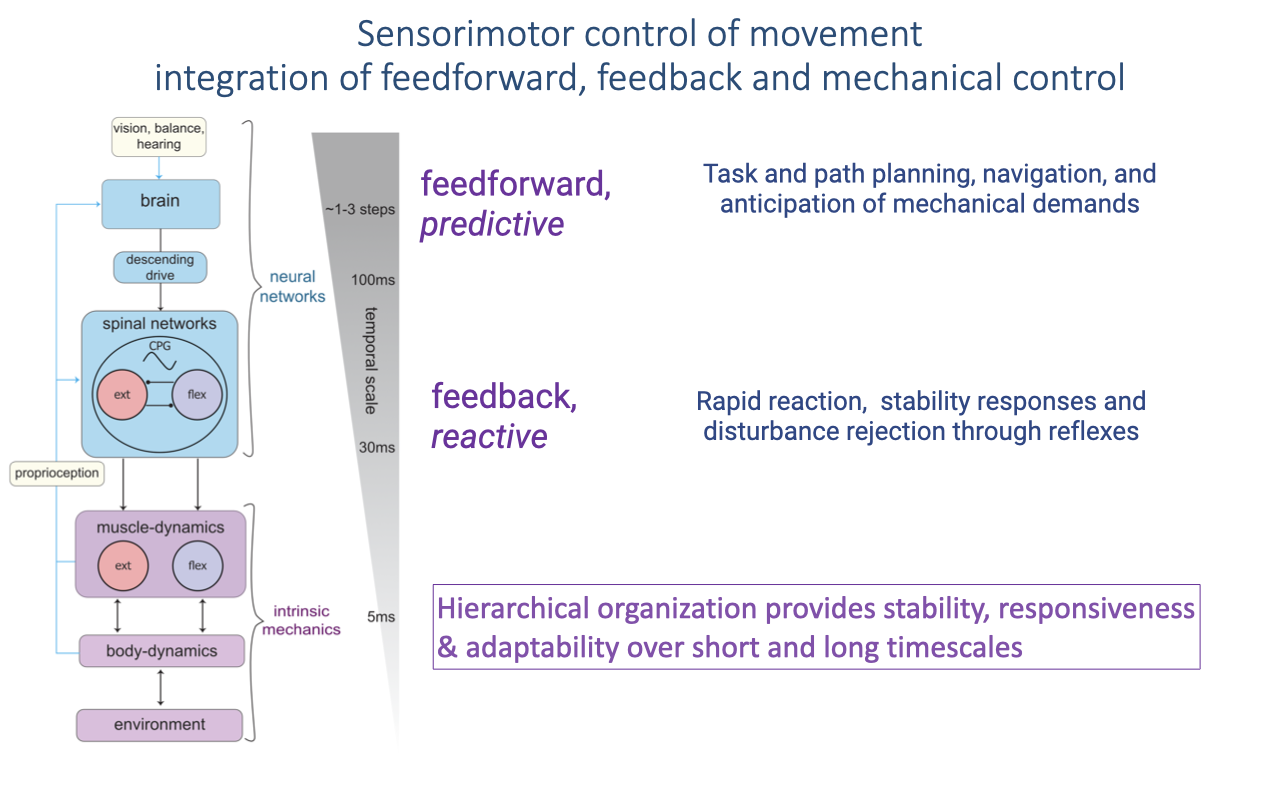
Sensorimotor Control of Movement
Locomotor control integrates feedforward (predictive) planning, feedback (reactive) reflexes, and intrinsic mechanical responses, organized hierarchically across timescales from ~5 ms to several steps (review Lecture 20).
This layered organization provides stability, responsiveness, and adaptability simultaneously.
Slide 32
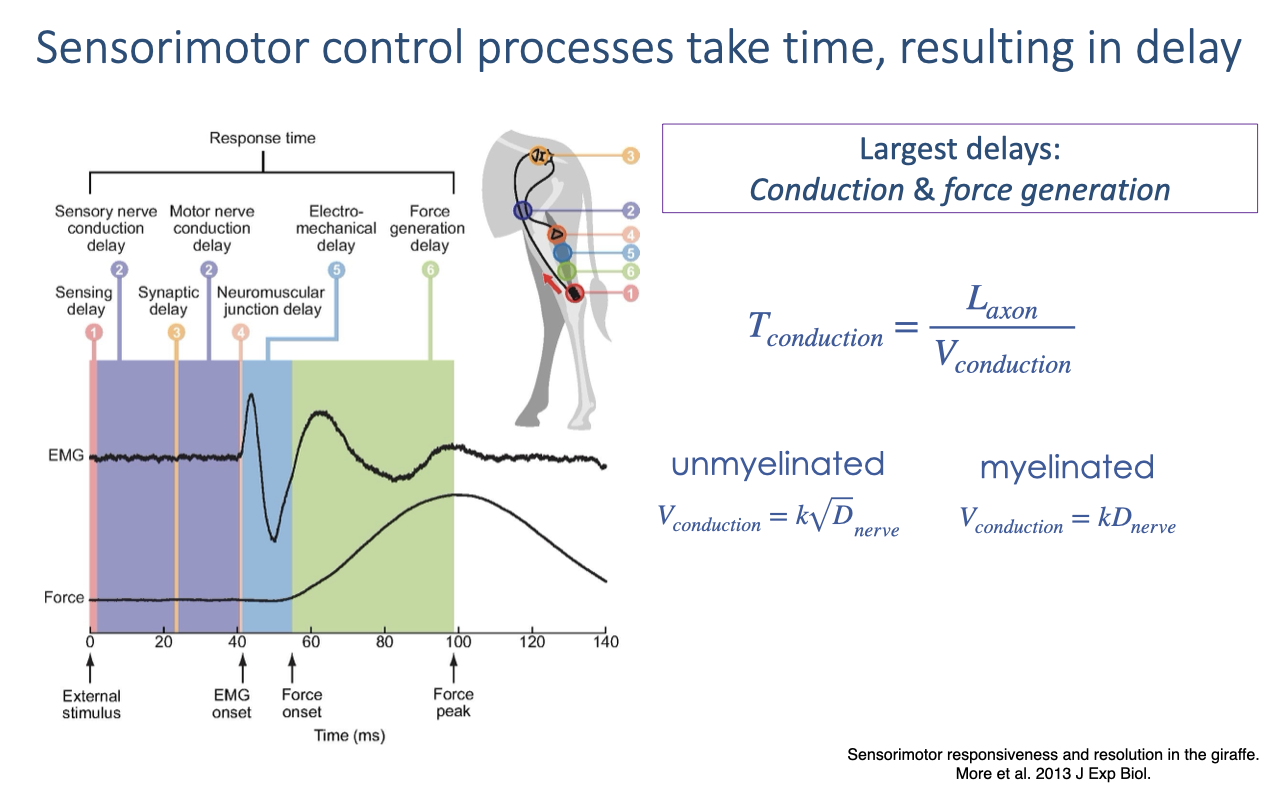
Sensorimotor Control Takes Time
Each control step adds delay — sensory and motor conduction, synaptic and neuromuscular-junction transmission, and electromechanical plus force-generation delay (review Lecture 20).
Conduction time depends on axon length and velocity (faster in larger and myelinated axons); the largest contributions are nerve conduction and force generation.
Slide 33
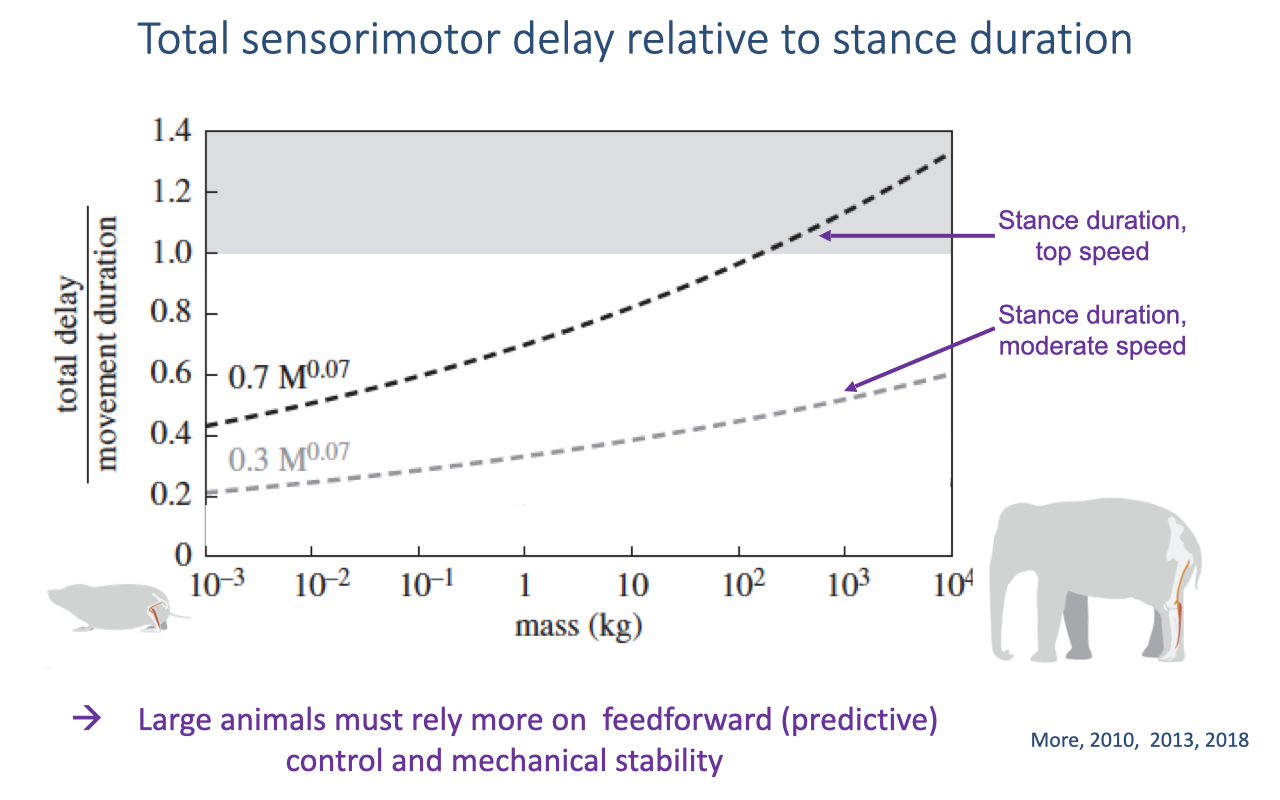
Total Sensorimotor Delay Relative to Stance Duration
Sensorimotor delay grows relative to stance duration as body size increases, so feedback corrections arrive too late within a step for the largest animals (review Lecture 20).
Large animals must therefore rely more on feedforward (predictive) control and intrinsic mechanical stability.
Humans fall on the larger side of this scaling, so at fast running speeds reflexes are too slow and movement must be planned predictively — a demand that may have contributed to the evolution of larger brains.
Slide 34
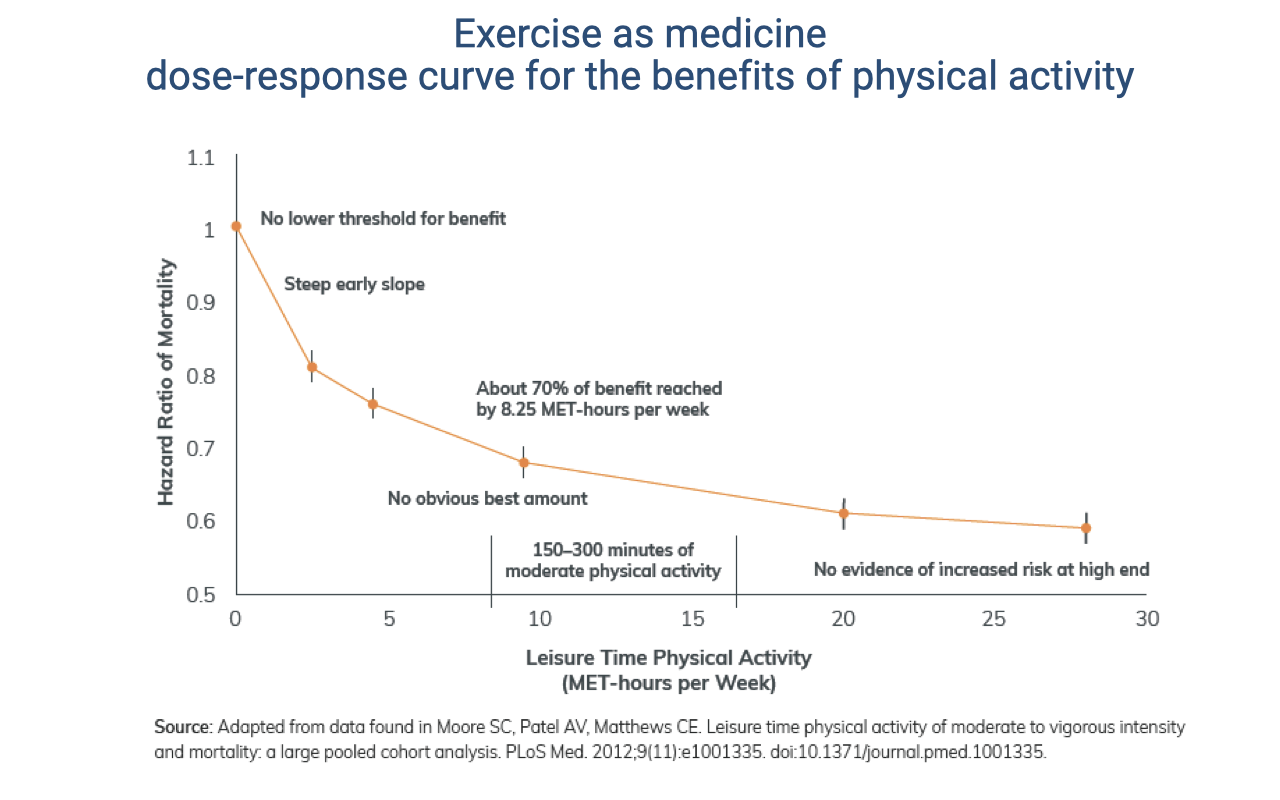
Exercise as Medicine: The Dose-Response Curve
Physical activity reduces mortality risk along a dose-response curve with no lower threshold and a steep early benefit — roughly 70% of the maximum benefit is reached near 8.25 MET-hours per week (review Lecture 21).
Benefits continue with more activity, with no clear optimal dose and no evidence of harm at the high end, assuming energetic needs are met and adequate rest avoids injury.
Across the population, higher activity is associated not only with lower mortality but also lower chronic-disease and cancer risk, better cancer survivability, and improved cognitive function and mood regulation.
Slide 35
The recap closes by inviting questions and encouraging students to revisit the lectures noted above to synthesize the course’s major themes — comparative and evolutionary context, the oxygen supply cascade, muscle structure and function across scales, the mechanics and energetics of locomotion, sensorimotor control, and exercise as medicine.